-
IntroducciónIntroducción al Curso
-
Sistema Inmunológico Sección 1Sistema inmunológico Innato1 Test
-
Sistema inmunológico Adquirido
-
Sobre las células T
-
Células B
-
Citoquinas
-
Células linfoides - Células asesinas naturales
-
Células Endoteliales
-
Respuesta inmune humoral
-
Respuesta inmune mediada por células
-
El microbioma, 70 por ciento de inmunidad
-
Células madre cancerosas


Selección clonal y diferenciación de células B
Las células B maduran en la médula ósea, donde se someten a una recombinación VDJ para producir receptores únicos que no reaccionan a los autoantígenos.
OBJETIVOS DE APRENDIZAJE
Describir la selección clonal y la diferenciación de células B
CONCLUSIONES CLAVE
Puntos clave
- En la médula ósea, la tolerancia central de las células B se produce mediante selección negativa. Las células B inmaduras se analizan para determinar su autorreactividad antes de abandonar la médula ósea. Las células B inmaduras cuyos receptores (BCR) se unen con demasiada fuerza a los antígenos propios no morirán.
- Cuando el receptor de células B en la superficie celular coincide con su antígeno afín en el cuerpo, las células B proliferan y secretan una forma libre de esos receptores (anticuerpos), con sitios de unión idénticos a los de la superficie celular original.
- Las células B que no han estado expuestas al antígeno, también conocidas como células B vírgenes, pueden activarse de forma independiente o dependiente de las células T.
- Hay dos tipos de activación independiente de células T: activación independiente de células T tipo 1 (policlonal) y activación independiente de células T tipo 2.
- Durante la selección clonal, las mutaciones aleatorias durante la expansión clonal provocan la producción de células B con mayor afinidad de unión a anticuerpos por sus antígenos.
- La hipótesis de la selección clonal puede explicar por qué las respuestas inmunitarias secundarias son tan efectivas para prevenir la reinfección por el mismo patógeno.
Términos clave
- Selección clonal : teoría de que los linfocitos portan receptores de antígenos antes de la activación y que las mutaciones aleatorias durante la expansión clonal provocan el desarrollo de linfocitos con alta afinidad de unión por sus antígenos.
- humoral : Perteneciente o relativo a los fluidos corporales o humores.
- Célula B de memoria : subtipo de células AB formado después de una infección primaria en la que la célula reconoce un epítopo específico.
Las células B son linfocitos que juegan un papel importante en la respuesta inmune humoral (a diferencia de la respuesta inmune mediada por células, que está gobernada por las células T). Las células B funcionan principalmente para producir anticuerpos contra antígenos, actúan como células presentadoras de antígenos (APC) y, finalmente, se convierten en células B de memoria para proporcionar inmunidad a largo plazo. Las células B se someten a una selección clonal y se desarrollan de manera similar a las células T con algunas diferencias notables.
Desarrollo de células B
Las células B inmaduras se producen en la médula ósea de la mayoría de los mamíferos. Su desarrollo ocurre a través de varias etapas, cada una de las cuales representa un cambio en el contenido del genoma en los loci de anticuerpos. Un anticuerpo está compuesto por dos cadenas ligeras (L) y dos pesadas (H) idénticas, y los genes que las especifican se encuentran en la región V (variable) y la región C (constante). La región V de cadena pesada tiene tres segmentos, V, D y J. Estos segmentos se recombinan aleatoriamente en un proceso llamado recombinación VDJ para producir un dominio variable único en la inmunoglobulina de cada célula B individual.
Se producen reordenamientos similares para la región V de la cadena ligera pero con solo dos segmentos involucrados: V y J. Cuando la célula B falla en cualquier paso del proceso de maduración, morirá por apoptosis, aquí llamada deleción clonal. Esta es una forma de selección positiva. Las células B también se prueban para determinar su autorreactividad mediante selección negativa. Si estas células B tienen una alta afinidad por unirse a los autoantígenos, morirán por deleción clonal u otra vía como la anergia.
Activación de células B
La activación de las células B se refiere a la diferenciación y expansión clonal de las células B. Cuando el receptor de las células B en la superficie celular coincide con su antígeno afín en el cuerpo, las células B proliferan y secretan una forma libre de esos receptores (anticuerpos) en el cuerpo. con sitios de unión idénticos a los de la superficie celular original. Sin embargo, el reconocimiento de antígenos por las células B no es el único elemento necesario para la activación de las células B. Las células B que no han estado expuestas al antígeno, también conocidas como células B vírgenes, pueden activarse de forma independiente o dependiente de las células T.
- La activación dependiente de las células T es la activación de las células B por las células T auxiliares tipo 2 en los ganglios linfáticos.
- La activación independiente de las células T ocurre cuando los antígenos se unen directamente a las propias células B, por lo general mediante la reticulación del antígeno con el receptor de la célula B o recibiendo el antígeno con un receptor de tipo toll.
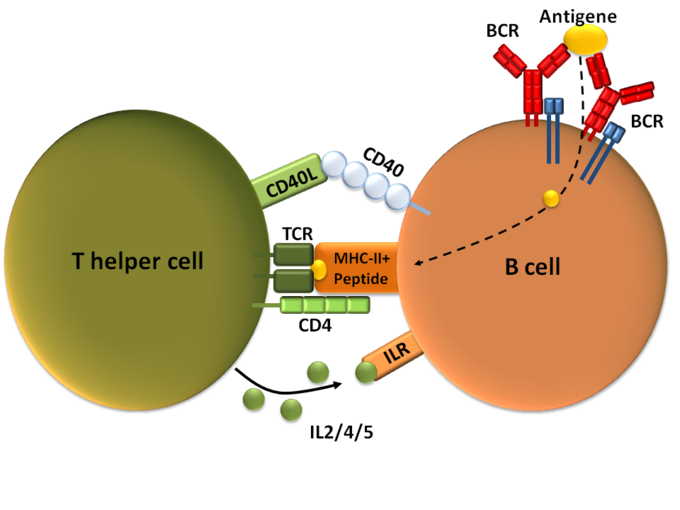
Activación de células B dependientes de T: activación de células B dependientes de células T, que muestra una célula TH2 (izquierda), una célula B (derecha) y varias moléculas de interacción.
Diferenciación de células B y expansión clonal
Después de la activación, la célula B sufre diferenciación y expansión clonal, que generalmente implica migración a centros germinales si la activación tiene lugar en un ganglio linfático. La diferenciación de células B es el proceso mediante el cual las células B se transforman en diferentes tipos, como células plasmáticas y blastos plasmáticos. La expansión clonal es el proceso por el cual las células hijas surgen de una célula madre. Durante la expansión clonal de células B, se producen muchas copias de esa célula B que comparten la afinidad y la especificidad del mismo antígeno.
Selección clonal
La selección clonal es una teoría que establece que las células B expresan receptores específicos de antígeno antes de que los antígenos se encuentren en el cuerpo. Después de la activación de las células B, las células B se clonan a sí mismas mediante expansión clonal, pero durante cada división celular, se producen mutaciones aleatorias que aumentan gradualmente la afinidad de unión de los anticuerpos producidos por las células B a los antígenos.
Por ejemplo, se cree que las células B de memoria que se diferencian después de una respuesta inmune adaptativa se someten a una selección clonal de modo que los anticuerpos producidos por las células B de memoria más nuevas tienen afinidades de unión considerablemente más altas con sus antígenos. Esta teoría puede explicar por qué las respuestas inmunitarias secundarias de las células de memoria son tan efectivas que las infecciones repetidas por el mismo patógeno se detienen incluso antes de que se desarrollen los síntomas. Después de la infección inicial, las mutaciones aleatorias durante la selección clonal podrían producir células B de memoria que pueden unirse más fácilmente a los antígenos que las células B originales.
Estructura y función de los anticuerpos
Un anticuerpo es una proteína en forma de Y producida por las células B para identificar y neutralizar antígenos en el cuerpo.
OBJETIVOS DE APRENDIZAJE
Describir la estructura y función de los anticuerpos.
CONCLUSIONES CLAVE
Puntos clave
- Un anticuerpo, también conocido como inmunoglobulina, es una proteína grande en forma de Y producida por las células B y utilizada por el sistema inmunológico para identificar y neutralizar objetos extraños como bacterias y virus.
- Cada punta de la “Y” de un anticuerpo contiene un paratopo (una estructura análoga a un candado) que es específico para un epítopo particular (análogamente análogo a una llave) en un antígeno, permitiendo que estas dos estructuras se unan con precisión.
- Aunque la estructura general de los anticuerpos es muy similar, una pequeña región en la punta de la proteína es extremadamente variable, lo que permite que existan millones de anticuerpos con diferentes sitios de unión al antígeno. Esta región se conoce como región hipervariable.
- Se encuentran cinco isotipos de anticuerpos en diferentes ubicaciones y realizan diferentes funciones específicas.
- La base de la Y juega un papel en la modulación de la actividad de las células inmunes. Esta región se denomina región Fc, y los fagocitos pueden unirse para iniciar la fagocitosis.
- Los anticuerpos que se unen a los antígenos de superficie de una bacteria atraen el primer componente de la cascada del complemento con su región Fc e inician la activación del sistema del complemento clásico.
Términos clave
- epítopo : parte de una biomolécula (como una proteína) que es el objetivo de una respuesta inmunitaria.
- paratope : Parte de la molécula de un anticuerpo que se une a un antígeno.
- isotipo : marcador correspondiente a un antígeno que se encuentra en todos los miembros de una subclase de una clase específica de inmunoglobulinas.
Un anticuerpo (formalmente llamado inmunoglobulina) es una gran glicoproteína en forma de Y producida por las células B y utilizada por el sistema inmunológico para identificar y neutralizar patógenos. Los anticuerpos son producidos por las células B y se secretan a la circulación o permanecen expresados en la superficie de la célula B.
Estructura de los anticuerpos
El anticuerpo reconoce una parte única de un antígeno (objeto extraño). Cada punta de la “Y” de un anticuerpo contiene un paratopo (una estructura análoga a un candado) que es específico para un epítopo particular (análogamente análogo a una llave) en un antígeno, permitiendo que estas dos estructuras se unan con precisión. Con este mecanismo de unión, un anticuerpo puede neutralizar su objetivo directamente o marcarlo para que lo ataquen otras partes del sistema inmunológico.

Anticuerpo : cada anticuerpo se une a un antígeno específico, una interacción similar a un candado y una llave.
Los anticuerpos son glicoproteínas que pertenecen a la superfamilia de las inmunoglobulinas, típicamente formadas por unidades estructurales básicas, cada una con dos cadenas pesadas grandes y dos cadenas ligeras pequeñas. La mayoría de los anticuerpos existen como monómeros, en los que tienen una sola subunidad en forma de “Y”, pero algunos anticuerpos pueden existir como dímeros (dos subunidades) o pentámeros (cinco subunidades). El paratopo se considera una región hipervariable y tiene la misma especificidad y afinidad de unión al antígeno que el receptor de células B de la célula B que creó el anticuerpo. En algunos isotipos, el extremo de la cola del anticuerpo se llama región constante y se aleja de los extremos paratobios en forma de Y, funcionando como una cola Fc a la que los fagocitos pueden unirse.
Isotipos
Cinco isotipos diferentes de anticuerpos desempeñan cada uno funciones diferentes y generalmente se encuentran en diferentes partes del cuerpo.
- IgA: un dímero secretado en las superficies de las mucosas, como el intestino, el tracto respiratorio y el tracto urogenital, que evita la invasión de la mucosa al cuerpo por patógenos. Es resistente a las enzimas proteolíticas que se encuentran en las mucosas gastrointestinales.
- IgD: funciona principalmente como un receptor de antígenos en las células B que no han sido expuestas a antígenos. Se ha demostrado que activa basófilos y mastocitos para producir factores antimicrobianos.
- IgE: se encuentra en la circulación y se une a los alérgenos, provocando la liberación de histamina de los mastocitos y basófilos. También protege contra gusanos parásitos.
- IgG: tiene cuatro formas diferentes y proporciona la mayor parte de la inmunidad basada en anticuerpos contra patógenos invasores como la mejor opsonina de cualquier tipo de anticuerpo. Esto se debe a que expresa una cola a la que se unen los receptores Fc de los fagocitos, lo que activa la fagocitosis. Es el único anticuerpo capaz de atravesar la placenta para dar inmunidad pasiva al feto y puede activar el sistema clásico del complemento.
- IgM: Expresada en la superficie de las células B (monómero) y en un pentámero secretado con muy alta avidez. Elimina patógenos en las primeras etapas de la inmunidad mediada por células B (humoral) antes de que haya suficiente IgG. Al igual que la IgG, también puede activar el sistema del complemento clásico.
Función de los anticuerpos
Los anticuerpos circulantes son producidos por células B clonales que responden específicamente a un solo antígeno. Los anticuerpos contribuyen a la inmunidad de tres formas: evitando que los patógenos entren o dañen las células al unirse a ellas (neutralización); estimular la eliminación de patógenos por macrófagos y otras células recubriendo el patógeno (opsonización); y desencadenando la destrucción de patógenos estimulando otras respuestas inmunes como la vía del complemento. El sistema del complemento inicia una larga cascada de producciones de proteínas que opsonizan un patógeno para la fagocitosis o lo lisan directamente formando un complejo de ataque a la membrana. Durante la opsonización, el anticuerpo expresa la cola de un receptor Fc en un macrófago, neutrófilo o célula asesina natural. La célula inmunitaria se unirá a la cola Fc del anticuerpo en lugar del patógeno en sí, lo que acelera el proceso de búsqueda de patógenos para fagocitar. Además, debido a que los anticuerpos tienen dos o más paratopos, a veces pueden unir patógenos, lo que hace que la fagocitosis sea más eficiente.
Papel del sistema de complemento en la inmunidad
El sistema del complemento es la capacidad de los anticuerpos y las células fagocíticas para eliminar patógenos de un organismo.
OBJETIVOS DE APRENDIZAJE
Describir el papel del sistema del complemento en la inmunidad.
CONCLUSIONES CLAVE
Puntos clave
- El sistema del complemento ayuda a los anticuerpos y las células fagocíticas a eliminar los patógenos de un organismo.
- El sistema del complemento consta de una serie de pequeñas proteínas producidas por la reacción de fase aguda en el hígado durante la inflamación.
- El sistema del complemento puede jugar un papel en enfermedades con un componente inmunológico y las del sistema nervioso central. La deficiencia de proteínas del complemento es una forma de inmunodeficiencia primaria.
- La ruta clásica del complemento comienza con la unión de anticuerpos, lo que provoca una reacción en cascada de las proteínas del complemento que forman gradualmente un complejo de ataque a la membrana.
- La vía alternativa del complemento suele ser estimulada por antígenos patógenos o toxinas en lugar de anticuerpos, y escinde C3 hasta que haya suficiente para continuar los pasos de la vía clásica del complemento desde la etapa de la convertasa C5.
- La vía de la lectina es homóloga a la vía clásica, pero con la opsonina, lectina de unión a manosa (MBL) y ficolinas, en lugar de C1 del anticuerpo. Esta vía usa proteasas en la MBL para formar C3 convertasa, que continúa los pasos de la vía clásica del complemento desde el paso de C3 convertasa.
- El sistema del complemento está regulado por proteínas de control del complemento, como la vía de aceleración de la descomposición, que evita que las proteínas del complemento formen MAC en las células del cuerpo.
Términos clave
- C5a : proteína del complemento que es un mediador inflamatorio de fase aguda, que causa vasodilatación y quimiotaxis de neutrófilos.
- complejo de ataque a la membrana : el complejo final de todas las vías del sistema del complemento que lisa el patógeno. Está compuesto por C5b, C6, C7, C8 y C9.
- lectina de unión a manano : proteína que se une a los carbohidratos de los patógenos para activar la vía del complemento de la lectina.
El sistema del complemento o “complementa” la capacidad de los anticuerpos y las células fagocíticas para eliminar los patógenos de un organismo. El sistema del complemento consta de una serie de pequeñas proteínas que se encuentran en la sangre, generalmente sintetizadas por el hígado como parte de la reacción de fase aguda durante la inflamación sistémica (a partir de la liberación de TNF-alfa). Cuando son estimuladas por uno de varios desencadenantes, las proteasas en el sistema escinden proteínas específicas para liberar citocinas e iniciar una cascada amplificadora de escisiones adicionales. El resultado final de esta cascada de activación es la amplificación masiva de la respuesta y activación del complejo de ataque de membrana que destruye las células (también llamado MAC). Hay tres vías diferentes por las que puede ocurrir el sistema del complemento.
Camino del complemento clásico
La vía clásica del complemento es la vía principal por la que se produce el sistema del complemento. Se compone de una cascada de muchos pasos con proteínas del complemento que se escinden entre sí en un orden secuencial:
- El anticuerpo se une a un antígeno en la superficie de un patógeno, activando la proteína del complemento C1.
- C1 actúa como una proteasa y escinde C2 y C4 para formar C4b2b.
- C42b convierte C3 en C3a y C3b, que forma una convertasa C5.
- La convertasa C5 escinde C5 en C5a y C5b.
- C5b forma un complejo con C6, C7 y luego C8 y C9, que se convierte en el complejo de ataque a la membrana que lisa el patógeno.
Tenga en cuenta que C5a tiene otras funciones en el sistema inmunológico, como causar vasodilatación durante la inflamación y estimular la quimiotaxis de neutrófilos. Además, las células del cuerpo expresan una glicoproteína llamada factor de aceleración de la descomposición, que descompone la convertasa C3 y C5 en las células del cuerpo. Este factor evita que se formen complejos de ataque de membrana en las células del cuerpo en condiciones normales.
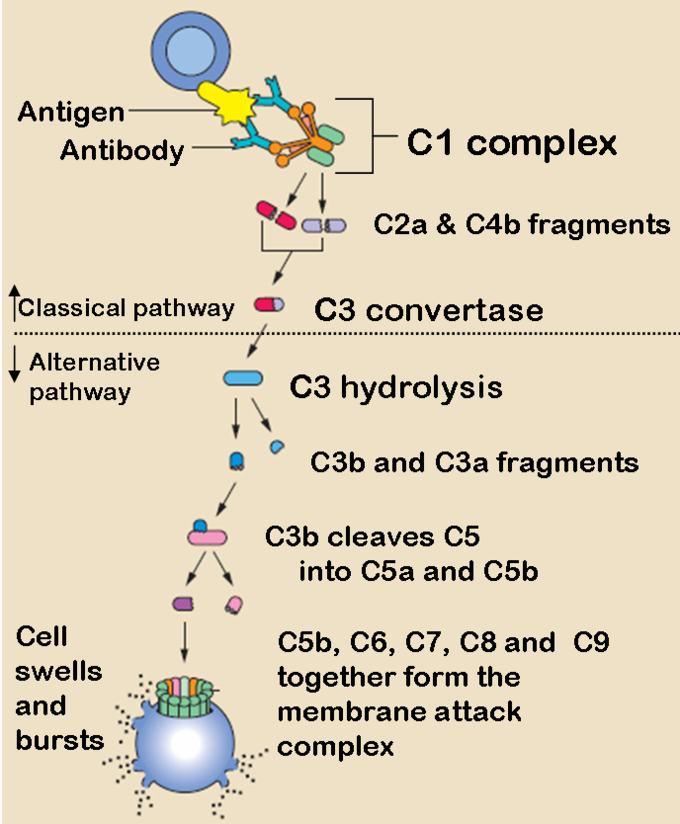
Las vías del complemento clásicas y alternativas: las vías del complemento clásicas y alternativas comienzan de manera diferente, pero terminan en la misma cascada de proteínas del complemento que se combinan para formar un complejo de ataque a la membrana.
La vía alternativa del complemento
La vía alternativa puede ser un precursor evolutivo sobrante de la vía clásica. A diferencia de la vía clásica, la vía alternativa generalmente es activada por mediadores inflamatorios microbianos en lugar de anticuerpos. Por ejemplo, el lipopolisacárido, la toxina de las bacterias gramnegativas, puede activar esta vía. Los pasos para la vía alternativa son:
- El antígeno patógeno (como LPS) activa C3, por lo que crea un complejo C3B
- El factor D escinde el complejo C3B para que se cree C3bBb.
- C3bBb es una convertasa C3, que convierte más C3 en C3a y C3b.
- De manera similar a la vía clásica, C3b forma un complejo C42b, y el resto de los pasos son esencialmente los mismos que en la vía clásica, terminando con C5b formando un complejo de ataque a la membrana con C6, C7, C8 y C9.
Vía de la lectina
La vía de la lectina no es causada por la unión de anticuerpos, sino por una proteína de unión a carbohidratos llamada lectina de unión a manano (MBL). Es un reactante de fase aguda producido en el hígado y se une a los carbohidratos en la superficie de muchos patógenos. Los pasos para la vía de la lectina son:
- MBL se une a los carbohidratos de un patógeno.
- Las proteasas unidas en el otro lado de la MBL escinden C4 en C4a y C4b.
- C4b crea C3 convertasa, y el resto de los pasos ocurren de manera idéntica a la ruta clásica desde el paso C3 convertasa.
Problemas con el sistema de complemento
El sistema del complemento puede desempeñar un papel en enfermedades con un componente inmunitario, como el síndrome de Barraquer-Simons, asma, lupus eritematoso, glomerulonefritis, diversas formas de artritis, cardiopatía autoinmune, esclerosis múltiple, enfermedad inflamatoria intestinal, lesiones por isquemia-reperfusión y rechazo de órganos trasplantados. El sistema del complemento también está cada vez más implicado en enfermedades del sistema nervioso central como la enfermedad de Alzheimer y otras afecciones neurodegenerativas como las lesiones de la médula espinal. Además, las deficiencias en las proteínas del complemento producidas en el hígado pueden conducir a una forma de inmunodeficiencia primaria (congénita), en la que el cuerpo es más susceptible a enfermedades, particularmente enfermedades autoinmunes e infecciones bacterianas graves.
Memoria inmunológica
La memoria inmunológica se refiere a la capacidad de las células B y T para producir células de memoria de larga duración que se defienden de patógenos específicos.
OBJETIVOS DE APRENDIZAJE
Describir la memoria inmunológica del sistema inmunológico.
CONCLUSIONES CLAVE
Puntos clave
- Cuando las células B y T comienzan a replicarse, algunos descendientes se convertirán en células de memoria de larga duración.
- Las células de memoria recuerdan todos los patógenos específicos encontrados durante la vida del animal y, por lo tanto, pueden provocar una fuerte respuesta si el patógeno vuelve a invadir el cuerpo.
- La inmunidad pasiva proviene de los anticuerpos IgG administrados a través de la madre durante el desarrollo fetal y a través de la leche materna. Esta memoria es a corto plazo, pero protege al bebé hasta que su propio sistema inmunológico adaptativo sea funcional.
- Durante una respuesta inmune secundaria, las células de memoria B y T trabajan para eliminar rápidamente el patógeno, evitando la reinfección por el mismo patógeno.
- Durante una vacuna, el antígeno de un patógeno se introduce en el cuerpo a través de una forma debilitada del patógeno que no puede causar una infección. Esto estimula al sistema inmunológico a desarrollar una inmunidad específica contra ese patógeno sin causar realmente la enfermedad que trae el patógeno.
- Las vacunas no existen para todos los patógenos debido a las frecuentes mutaciones de cepas y los desafíos para producir una inmunización lo suficientemente fuerte como para funcionar, pero no lo suficientemente fuerte como para causar una infección.
Términos clave
- respuesta inmune secundaria : El acto de exposición al mismo patógeno después de la respuesta inmune inicial. Las células de memoria B y T trabajan para eliminar rápidamente el patógeno para prevenir la reinfección.
- Vacunación : Inoculación con la forma debilitada de un patógeno para proteger contra una enfermedad o cepa de enfermedad en particular estimulando el desarrollo de la memoria inmunológica contra ese patógeno.
Cuando las células B y T comienzan a replicarse durante una respuesta inmune adaptativa, algunos descendientes se convierten en células de memoria de larga duración. Estas células de memoria recuerdan todos los patógenos específicos encontrados durante la vida del animal y, por lo tanto, pueden provocar una respuesta más fuerte, llamada respuesta inmune secundaria, si el patógeno invade el cuerpo nuevamente. El sistema inmunológico adaptativo se llama así porque es el resultado de una adaptación a una infección. La memoria inmunológica puede existir en la memoria activa a largo plazo o en la memoria pasiva a corto plazo.

Respuesta inmune : cuando las células B y T comienzan a replicarse, parte de la descendencia que producen terminará convirtiéndose en células de memoria de larga duración. Estas células de memoria recordarán todos los patógenos específicos encontrados durante la vida del animal y, por lo tanto, pueden provocar una fuerte respuesta si el patógeno vuelve a invadir el cuerpo.
Memoria pasiva
Los bebés recién nacidos son particularmente vulnerables a las infecciones ya que no han tenido una exposición previa a patógenos. Por tanto, la madre protege al bebé mediante varias capas de protección pasiva. Durante el embarazo, la IgG, un determinado isotipo de anticuerpo, se transporta al bebé desde la madre a través de la placenta, por lo que incluso los bebés tienen altos niveles de anticuerpos con especificidades antigénicas similares a las de la madre. Incluso la leche materna contiene anticuerpos que se transfieren al tracto gastrointestinal del bebé y protegen contra las infecciones bacterianas hasta que el bebé es capaz de producir sus propios anticuerpos. Dado que el feto no produce células de memoria ni anticuerpos, esto se denomina inmunidad pasiva. La inmunidad pasiva es de corta duración, desde un par de días hasta un par de meses.
A medida que el bebé madura, su timo y médula ósea trabajan para generar una reserva de linfocitos maduros que forman la base del sistema inmunológico adaptativo personal del bebé. Debido a que la memoria pasiva proviene de los anticuerpos en lugar de las propias células B, los bebés no heredan la memoria inmunológica a largo plazo de la madre. Incluso si el bebé recibe anticuerpos específicos para ciertas enfermedades de su madre, el bebé no podría reforzar una memoria a largo plazo que dirigiría la exposición y presentación del antígeno.
Inmunización y memoria activa
Después de una infección, la memoria activa a largo plazo se adquiere mediante la activación de las células B y T. Las células de memoria se derivan de sus células B y T parentales y se someten a una selección clonal después de la infección, lo que aumenta la afinidad de unión al antígeno. Después de la reinfección, la respuesta inmunitaria secundaria normalmente elimina el patógeno antes de que aparezcan los síntomas de una infección. Durante la respuesta inmune secundaria, las células T de memoria proliferan rápidamente en células T cooperadoras activas y citotóxicas específicas para ese antígeno, mientras que las células B de memoria producen rápidamente anticuerpos para neutralizar el patógeno. La memoria activa a largo plazo consiste en una respuesta rápida y forma una memoria inmunológica permanente mientras esas células de memoria sobreviven.
Las vacunas aprovechan el desarrollo de linfocitos de memoria al generar inmunidad activa artificialmente, un proceso llamado inmunización. Durante una vacunación, el antígeno de un patógeno se introduce en el cuerpo y estimula al sistema inmunológico a desarrollar una inmunidad específica contra ese patógeno. No causa la enfermedad que trae el patógeno porque la vacuna usa una forma atenuada del patógeno que contiene el mismo antígeno pero no tiene la capacidad de replicación. Esta introducción deliberada del patógeno tiene éxito ya que explota la especificidad e inducibilidad naturales del sistema inmunológico. La vacunación es una manipulación extremadamente eficaz del sistema inmunológico que ayuda a combatir enfermedades. A lo largo del desarrollo de la vacuna, han salvado innumerables vidas,
A pesar de la eficacia de las vacunas, todavía no existen métodos para desarrollar vacunas para todos los patógenos. Muchos patógenos sufren mutaciones que cambian la expresión de sus antígenos, haciendo infructuosos los intentos de inmunización para enfermedades como el resfriado común o el norovirus. Muchos patógenos parasitarios, como el plasmodium protist que causa la malaria, no se han vacunado con éxito porque es un desafío desarrollar una vacuna que sea lo suficientemente fuerte como para estimular una respuesta inmune (inmunogenicidad suficiente) sin causar una infección viva.
Antígenos del complejo mayor de histocompatibilidad (autoantígenos)
El complejo principal de histocompatibilidad (MHC) es una molécula de la superficie celular que regula las interacciones entre los glóbulos blancos y otras células.
OBJETIVOS DE APRENDIZAJE
Describir los principales antígenos del complejo de histocompatibilidad.
CONCLUSIONES CLAVE
Puntos clave
- El complejo mayor de histocompatibilidad (MHC) es una molécula de superficie celular que media las interacciones entre los glóbulos blancos y otros leucocitos o células.
- El MHC se divide en subgrupos de clase I, clase II y clase III, todos codificados por el mismo gen.
- La presentación del antígeno MHC gana su diversidad a partir del alto grado de polimorfismo de la propia proteína MHC.
- La diferencia de HLA entre miembros de la misma especie es la principal razón por la que el receptor del órgano rechaza los órganos trasplantados.
Términos clave
- antígeno leucocitario humano (HLA) : nombre del complejo principal de histocompatibilidad (MHC) en humanos.
El complejo mayor de histocompatibilidad (MHC) es una molécula de la superficie celular codificada por una gran familia de genes en todos los vertebrados. Las moléculas de MHC median las interacciones de los leucocitos con otros leucocitos o células corporales. El MHC determina la compatibilidad de los donantes para el trasplante de órganos, así como la susceptibilidad de uno a una enfermedad autoinmune a través de la inmunización de reacción cruzada. En los seres humanos, el MHC también se denomina antígeno leucocitario humano (HLA), porque el MHC a menudo puede actuar como un antígeno para los leucocitos humanos.
Tipos de MHC
MHC es una proteína polimórfica adherida a la superficie de las células. Tres clases diferentes de MHC difieren en estructura y función en el sistema inmunológico, pero todas están codificadas por el mismo conjunto de secuencias de genes.
- MHC I se presenta en todas las células del cuerpo. Contiene un epítopo que forma el sitio de unión estructural de un antígeno. El MHC I interactúa con las células asesinas naturales (NK) y las células T citotóxicas para indicar si una célula es propia o no, y si contiene un antígeno específico de esa célula T.
- El MHC II se presenta principalmente en macrófagos, células dendríticas y linfocitos T auxiliares, todos involucrados en la presentación de antígenos. Tiene una región helicoidal más larga que el MHC I, lo que le permite unirse a CD4 (células T colaboradoras) durante la presentación del antígeno.
- MHC III es una enzima secretada que no está unida a la membrana ni participa en la presentación de antígenos como MHC I y II. Simplemente se incluye como una proteína MHC porque está codificada por el mismo conjunto de genes. Participa en la producción de proteínas del complemento y citocinas inflamatorias.
Durante el procesamiento de antígenos antes de la presentación, los péptidos proteicos de patógenos y MHC (I o II) viajan a través del citoplasma y luego a la superficie de la membrana celular. Una serie compleja de vesículas permite el transporte de MHC desde la célula, y el retículo endoplásmico y los cuerpos de Golgi facilitan este transporte durante el procesamiento del antígeno. Después del procesamiento, el MHC puede presentar el antígeno peptídico unido a él a las células T vírgenes.